Abstract
The debate on human pheromones has been an ongoing topic in research. While commonly cited human pheromones (androstenone, androstenol, androstadienone, estratetraenol) lack conclusive scientific validation due to methodological flaws and inconsistent evidence, many studies still identify a positive relationship between some human odorants and human behavior. This paper provides a critical overview of research on these ‘putative human pheromones’ by conducting an extensive investigation into the molecular misstatements in the methodologies of supporting studies. The research suggests that the most probable precursors of human pheromones are carboxylic acids and sulfur compounds. The main issue in testing any possible odors is their concentration, which is often disregarded and can only be accurately determined through rigorous bioassay. Only when a specific concentration that can impact behavior or physiology is established, can the reduced response to pheromones offer insights into their significance in modern social interactions.
Introduction
Pheromones are chemical cues used in communication within one species. They can be mainly split into two categories: releasers, that shape behavior changes, and primers, that influence development process by neuroendocrine responses. At the molecular level, pheromones can be structurally diverse and often consist of species-specific combinations of molecules rather than single compounds. These molecules are typically volatile, allowing them to be easily dispersed and detected by other members of the same species.
Pheromones have been discovered across the entire animal kingdom. One of the first observations in mammals came in the late 19th century when Darwin studied the breeding seasons of male goats. Pheromones play various roles depending on the species, including attracting mates, influencing mate selection, guiding trail following, and facilitating interactions between parents and offspring [2]. Nevertheless, a key aspect of pheromones is that, regardless of their specific function, the reactions they provoke appear to be “innate” within the species, despite individual variations in experience or context.
From an evolutionary perspective, it is highly likely that humans possess pheromones. Like other primates, we develop odor-producing secretions from apocrine glands during puberty, which accompany sexual maturation. This suggests that, although our responses to pheromones may have weakened due to a lack of selection pressure, we likely still secrete them, like our closest relatives, the great apes. While great apes, like humans, don’t rely as heavily on odor communication as other primates, they still have secretory glands that could produce pheromones, indicating a continued role for odors in our biology [12]. Moreover, relatively new findings claim that despite not having functional vomeronasal system believed to be specialized in pheromones detection, our main olfactory system should also enable us their recognition [8, 13]
Based on the above, over the past 50 years, many researchers have supported the existence of human pheromones through molecular findings; however, none of these studies have yet reached full scientific validity. The molecules in question include four androstene steroids: androstenone, androstenol, androstadienone, and estratetraenol. These compounds, often referred to as ‘putative human pheromones,’ were first presented by Monti-Bloch & Grosser at a 1991 conference sponsored by the EROX Corporation, which had commercial interests in human pheromones. While they suggested these molecules could function as pheromones, no details were provided on their extraction, identification, or bioassay, leaving no information to illustrate how these molecules were identified. The claims were built on the work of Jacob & McClintock, who cautioned that it was premature to label these steroids as human pheromones.
To address these ongoing debates and gaps in research, this paper aims to critically evaluate the current evidence supporting the existence and function of human pheromones. By examining recent studies, methodological approaches, and the molecular basis of these compounds, the objective is to provide a clearer understanding of whether these purported human pheromones play a significant role in human behavior and communication, and how their molecular mechanisms contribute to their potential effects.
Business Management
In-person
Cultivate leadership skills in New York, where multinational...

Coding
In-person
Master the language of technology in Cambridge, a...

Criticism around common “putative human pheromones”
The four the most often called “human pheromones” include: androstenone (5α-androst-16-en-3-one), androstenol (5α-androst-16-en-3α-ol), androstadienone (Δ4,16-androstadiene-3-one) and estratetraenol (estra-1, 3,5 (10),16-tetraen-3-ol). According to Wyatt (2014), these molecules have been studied mostly in two waves. Between 1970-1990s, androstenone and androstenol were used as key molecules in pheromone research, but from 1991 and especially after 2000, androstadienone and estratetraenol emerged as the primary ‘putative human pheromones,’ based on unpublished identifications by a US corporation.
For independent onlookers or enthusiasts of science, the results corroborating pheromones existence seem highly probable as their molecular basis prompting steroid character is linked to other mammalian pheromones. Scientists seem to have experimentally proven their presence in human fluids like apocrine sweat or saliva, and what is more intriguing, different sensitivity to their concentration between female sand males [4]. Additionally, there is evidence of varying sensitivity to these compounds between males and females, and over 40 papers have reported physiological or psychological effects linked to these “putative human pheromones” since 2000 [2]. Although obtaining positive results, they have no scientific basis that could demonstrate the studied molecules as human pheromones.
Critics argue that positive results may be compromised by methodological flaws, such as small sample sizes, statistical errors, and publication bias [1]. For instance, the very low concentrations of androstenone and related steroids in axillary sweat limit generalizability, and many individuals cannot even detect these compounds. In fact, a study by Nixon et al. (1988) found that only 10 out of 24 males had detectable levels of these steroids in their sweat. On the other hand, some studies face the lack of peer review and a paucity of cause-and-effect clarification, which may be mistaken for effects generated by non-pheromone odors that impact human mood. This has created a self-reinforcing cycle in which the popularity of androstadienone and estratetraenol in research, despite limited evidence, leads to studies and citations building upon each other, thereby perpetuating the belief in these molecules as pheromones without rigorous validation [2].
Join the Immerse Education 2025 Essay Competition
Follow the instructions to write and submit your best essay for a chance to be awarded a 100% scholarship.

Significant discoveries using other approaches
However, not all experiments are utterly unhelpful in the search for molecular bases of human attraction. For example, Wedekind’s experiment with smelly t-shirts, where female participants rated the attractiveness of the shirts, is globally recognized as influential in evolutionary psychology. Although the term “pheromones” is not used in Wedekind’s original paper, his research demonstrates that smells, particularly those related to the major histocompatibility complex (MHC), play a significant role in mate selection by influencing preferences based on genetic compatibility [19].
That is why many researchers continue to research pheromones by choosing different approaches, for example comparative anatomical studies. In 2009, using this approach scientists found out that the the humans’ vomeronasal organ (VNO), also known as Jacobson’s organ, compared to other animals’ one is poorly developed [8]. While some people have a small structure resembling the VNO, it appears to lack the necessary neural connections to the brain and the specific receptors found in other mammals that are essential for pheromone detection. However, despite traditional view, evidence suggests that both vomeronasal and main olfactory systems are involved in pheromonal communication, and not only Jacobson’s organ. Thus, it can be concluded that humans even relying solely on the main olfactory system can perhaps detect pheromones. Furthermore, the pheromone molecules can act as ligands, binding themselves to specific receptors through a hydrophobic pocket, on the surface of sensory neurons in the vomeronasal organ or the main olfactory epithelium [20]. These receptors, such as V1Rs, V2Rs, and formyl peptide receptors (FPRs), are G protein-coupled receptors (GPCRs) specialized for pheromone detection. The binding of ligands to these receptors triggers intracellular signaling pathways, generating electrical signals that are sent to the brain, where they are processed to elicit behavioral responses.
Potential pheromones contributing to human axillary odor
The precursors for potential pheromones in human axillary odor are metabolized locally and later secreted from axillary glands as odorless conjugates that are later converted into volatile odorants by bacterial enzymes. These precursors include carboxylic acids, such as 3-methyl-2-hexenoic acid (3M2H) and 3-hydroxy-3-methyl-hexanoic acid (HMHA), which are linked to a glutamine residue; sulfur compounds, released as odorant sulfur compounds from a cysteine–glycine dipeptide; and steroids like 5-α-androst-16-en-3α-ol, detected as a glucuronide conjugate in axillary secretions [9].
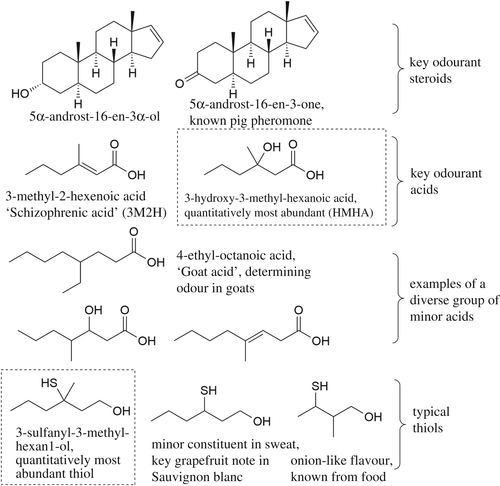
Figure 1. The structure of the known key human axilla odorants [9]
Key odorous compounds, such as short, straight-chain fatty acids (e.g., hexanoic acid, octanoic acid, isovaleric acid), are present in sweat, but they are not the primary contributors to the characteristic axilla odor in sexually mature individuals with active apocrine glands [9]. Although initially believed that incomplete β-oxidation of long-chain fatty acids was the main cause of odor, this theory has been refuted in favor of the idea that the characteristic axillary scent is primarily linked to amino acid degradation products from bacterial breakdown of secreted proteins and dead keratinocytes. Therefore, the role of microbiome has become more notable as it synthesis these precursors into volatile fatty acids and other odoriferous compounds via enzymatic processes like hydrolysis, oxidation and fermentation.
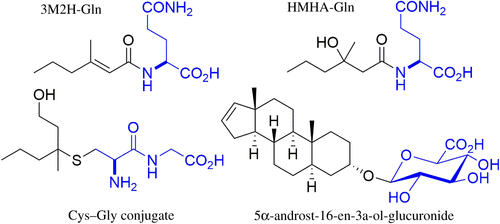
Figure 2. The structure of the three classes of odorants precursors [9]
Firstly, after being locally metabolized, odorless conjugates—such as carboxylic acids, sulfur compounds, and steroids—are transported into secretory vesicles via ABCC11, where they undergo processing by enzymes like γ-glutamyl transferase 1 (GGT1), dipeptidase, and β-lyase [9]. These enzymatic actions lead to the formation of a Cys-Gly conjugate, which is then cleaved by bacteria on the skin surface, releasing volatile sulfur compounds. Key functional groups, such as the thiol group in the GSH conjugate, play a crucial role in forming these volatile compounds, while amide groups in the glutamine side chain enhance enzyme specificity through hydrogen bonding. Once released, these low-molecular-weight thiols passively diffuse through the bacterial membrane and evaporate from the skin, forming the basis of axillary odor. The enzymes involved in this process are characterized by metal-binding domains containing zinc atoms essential for catalytic activity, with active sites featuring hydrophobic pockets that allow substrate flexibility.
Troublesome methodology
One of the main issues with previously mentioned studies advocating for the existence of “putative human pheromones” is their lack of suitable methodology. According to Wyatt (2014) the most reliable technique could be bioassay, an experimental method that helps tracking the effect of a substance to eventually determine the concentration at which a biology activity can take place. This way the established results could act as the minimum concentration at which the speculated pheromone could have any impact on a person, as it was in the case of bombykol, the first identified insect pheromone.
To accomplish that scientists would have to follow several steps just as Adolf Butenandt in 1959, when he used bioassay techniques to measure the attraction of male silkworm moths to the chemical released by females. The initial step requires observation of behavioral or physiological response that is hypothesized to be linked with a specific secreted chemical substance. Then, the bioactive molecules would have to be isolated, chemically identified and synthesized. The latter is essential to confirm that the proposed molecules, at natural concentrations, are both necessary and sufficient to replicate the original activity when tested with the initial bioassay.
Moreover, we should first stop using androstadienone and estratetraenol as stimuli, as there’s no solid evidence they are ‘human pheromones,’ despite their common experimental use. To identify new molecules secreted among hundreds or thousands of other compounds from axillary areas, a subtractive analysis can be more effective. This approach involves directly comparing results from two or more groups of humans that differ by a specific trait, similar to how 4-ethyloctanal was identified as a key pheromone in goats despite its low concentration relative to other molecules [16]. Consequently, a promising next step in human pheromone research could involve comparing the volatile compounds emitted by adult males, females, and prepubescent children, with a focus on identifying molecules that appear with the maturation of sebaceous and apocrine glands during puberty [2].
Such study of volatile pheromones highlights the need for accurate, efficient, and non-invasive methods compared to traditional approaches like collecting odors on cotton pads. On of the primary techniques include Gas Chromatography-Mass Spectrometry (GC-MS) that separates volatile compounds through gas chromatography (GC) based on their boiling points and then identifies them via mass spectrometry (MS), which provides a molecular “fingerprint” of each compound. This method has been already instrumental in identifying key organoleptic contributors in axillary secretions [15]. Another promising approach involves identifying ligands for highly conserved human olfactory receptors (ORs), which are potentially under strong stabilizing selection due to their role in pheromone detection, though this remains largely theoretical due to limited knowledge of ligands for human ORs [18]. Furthermore, calcium imaging is a crucial technique used to study pheromone receptors by observing changes in calcium levels in response to pheromone binding, allowing researchers to track these ligand-receptor interactions in real-time [8].
Discussion on evolution
Given the speculative roles of pheromones in mate selection, kin recognition, social bonding, and health detection through axillary odor, understanding the function of the vomeronasal organ in early humans is increasingly important for the debate on the evolution of chemical communication. The dormant state of the vomeronasal organ (VNO) sheds light on how humans have become less reliant on axillary odors for social interactions and mate selection. This adaptation is evident in the ABCC11 gene mutation, particularly prevalent in East Asian populations, which affects the production of odor precursors in sweat. Cultural factors may have influenced this evolutionary shift, leading to a diminished role of body odor in social dynamics [10]. Consequently, individuals with this mutation may not have faced significant disadvantages in social interactions or mate selection, reflecting a potential loss of the adaptive advantage once provided by body odor.
It cannot be omitted that humans’ responses to odors are often subtle and often context-dependent. Therefore, to prove such evolutionary theories about olfactory communication, it is pivotal to examine less culturally influenced populations, such as infants. For instance, Doucet and Montgomery’s research on infant and maternal odor detection demonstrates that newborns exhibit an instinctive response to the odors secreted by any lactating mother, not just their own, highlighting the fundamental role of olfactory cues in early bonding. Thus, understanding these primal interactions can provide insights into the evolutionary significance of olfactory communication.
Conclusion
Despite extensive research, progress in establishing the existence of human pheromones remains limited, primarily due to methodological challenges and inconsistent findings. Although there is evidence suggesting the presence of potential human pheromones, scientific validation is incomplete. Addressing these gaps requires rigorous bioassays and advanced chemical analysis techniques, emphasizing the need to study natural behaviors and avoid assumptions based on cross-species data. However, promising avenues for future research include the identification and analysis of specific molecules, such as carboxylic acids and sulfur compounds, using refined methodologies emerges. Additionally, the evolution of human olfactory communication—evidenced by changes in the vomeronasal organ and cultural shifts in body odor perception—indicates a reduced role for pheromones in modern social interactions. By focusing on less culturally influenced populations, such as infants, researchers can gain clearer insights into the molecular basis of pheromones and their evolutionary significance in human behavior.
Bibliography
1. Wyatt TD. The search for human pheromones: the lost decades and the necessity of returning to first principles. Proceedings of the Royal Society B: Biological Sciences. 2015 Apr 7;282(1804):20142994.
2. Wyatt TD. Pheromones and animal behavior : chemical signals and signatures. Cambridge, United Kingdom: Cambridge University Press; 2014.
3. Wikipedia Contributors. Richard L. Doty [Internet]. Wikipedia. Wikimedia Foundation; 2023 [cited 2024 Aug 21]. Available from: https://en.wikipedia.org/wiki/Richard_L._Doty
4.Nixon A, Mallet AI, Gower DB. Simultaneous quantification of five odorous steroids (16-androstenes) in the axillary hair of men. Journal of Steroid Biochemistry. 1988 May 1;29(5):505–10.
5. Wyatt TD. Reproducible research into human chemical communication by cues and pheromones: learning from psychology’s renaissance. Philosophical Transactions of the Royal Society B: Biological Sciences. 2020 Apr 20;375(1800):20190262.
6. Monti-Bloch L, Grosser BI. Effect of putative pheromones on the electrical activity of the human vomeronasal organ and olfactory epithelium. The Journal of Steroid Biochemistry and Molecular Biology [Internet]. 1991 Oct 1;39(4, Part 2):573–82. Available from: https://www.sciencedirect.com/science/article/abs/pii/0960076091902554
7. Jacob S, McClintock MK. Psychological State and Mood Effects of Steroidal Chemosignals in Women and Men. Hormones and Behavior. 2000 Feb;37(1):57–78.
8. Tirindelli R, Dibattista M, Pifferi S, Menini A. From Pheromones to Behavior. Physiological Reviews. 2009 Jul;89(3):921–56.
9. Natsch A, Emter R. The specific biochemistry of human axilla odour formation viewed in an evolutionary context. Philosophical Transactions of the Royal Society of London Series B, Biological Sciences [Internet]. 2020 Jun 8;375(1800):20190269. Available from: https://pubmed.ncbi.nlm.nih.gov/32306870/
10. Yoshida K, Yamashita H, Conroy S, Marks M, Kumar C. A Japanese version of Mother-to-Infant Bonding Scale: factor structure, longitudinal changes and links with maternal mood during the early postnatal period in Japanese mothers. Archives of Women’s Mental Health. 2012 Jun 26;15(5):343–52.
11. Robinson C. The Biochemistry of Attraction. Journal of Student Research at Indiana University East [Internet]. 2021 Jun 13;3(1):126–39. Available from: https://scholarworks.iu.edu/journals/index.php/jsriue/article/view/31822/36526
12. Stevenson RJ. An Initial Evaluation of the Functions of Human Olfaction. Chemical Senses [Internet]. 2010 Jan 1;35(1):3–20. Available from: https://academic.oup.com/chemse/article/35/1/3/305000
13. Baxi KN, Dorries KM, Eisthen HL. Is the vomeronasal system really specialized for detecting pheromones? Trends in Neurosciences. 2006 Jan;29(1):1–7.
14. Gower DB, Holland KT, Mallet AI, Rennie PJ, Watkins WJ. Comparison of 16-Androstene steroid concentrations in sterile apocrine sweat and axillary secretions: Interconversions of 16-Androstenes by the axillary microflora—a mechanism for axillary odour production in man? The Journal of Steroid Biochemistry and Molecular Biology. 1994 Mar;48(4):409–18.
15. Cotton S. In pursuit of Bombykol [Internet]. RSC Education. 2000. Available from: https://edu.rsc.org/feature/in-pursuit-of-bombykol/2020169.article
16. Murata K, Tamogami S, Itou M, Ohkubo Y, Wakabayashi Y, Watanabe H, et al. Identification of an Olfactory Signal Molecule that Activates the Central Regulator of Reproduction in Goats. Current Biology. 2014 Mar;24(6):681–6.
17. Penn DJ, Oberzaucher E, Grammer K, Fischer G, Soini HA, Wiesler D, et al. Individual and gender fingerprints in human body odour. Journal of The Royal Society Interface. 2006 Nov 28;4(13):331–40.
18. Peterlin Z, Firestein S, Rogers ME. The state of the art of odorant receptor deorphanization: A report from the orphanage. Journal of General Physiology. 2014 Apr 14;143(5):527–42.
19. Wedekind C, Seebeck T, Bettens F, Paepke A. MHC-Dependent Mate Preferences in Humans. Proceedings: Biological Sciences [Internet]. 1995 [cited 2024 Aug 21];260:245–9. Available from: https://www.coherer.org/pub/mhc.pdf
20. Malnic B. Searching for the Ligands of Odorant Receptors. Molecular Neurobiology. 2007 Jun 19;35(2):175–81.
21. Doucet S, Soussignan R, Sagot P, Schaal B. The Secretion of Areolar (Montgomery’s) Glands from Lactating Women Elicits Selective, Unconditional Responses in Neonates. Hausberger M, editor. PLoS ONE. 2009 Oct 23;4(10):e7579.